
- Product Name
- CasNo
- MF
- MW
- Content
- Appearance
- Packing
- Apply
- 1,4-Diaminobutane
- 110-60-1
- C4H12N2
- 88.1527
- colourless liquid
Your Location:Home > Products > Catalysts and additives > 1,4-Diaminobutane
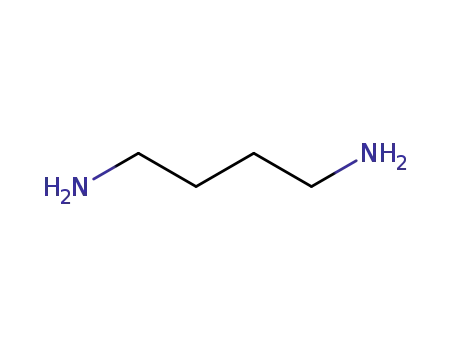
|
Biosynthesis |
l ,4-Diaminobutane (putrescine), spermidine, spermine, and closely related derivatives are found in a wide variety of animals, bacteria, yeasts, and plants. It is generally accepted that polyamine biosynthesis is intimately interrelated with the synthesis of nucleic acids and proteins[3]. Polyamines are ubiquitous in biological materials, although the relative amounts of 1,4-diaminobutane, spermidine, and spermine differ markedly in different cells[4, 5]. In general, prokaryotes have a higher concentration of 1,4-diaminobutane than spermidine and lack spermine. Eukaryotes usually have little 1,4-diaminobutane, and have spermine as well as spermidine. The pathway for the biosynthesis of 1,4-diaminobutane and spermidine was first established in microorganisms and was later found to be very similar in animal cells. In bacteria, 1,4-diaminobutane may be formed either by ornithine decarboxylase or by arginine decarboxylase via agmatine[6-8]. Both biosynthetic enzymes are normally present in Escherichia coli, although ornithine decarboxylation is usually the major pathway[8]. Both decarboxylases have been purified from E. coli and have been shown to require pyridoxal phosphate[9-11]. These enzymes are subject to feedback inhibition and repression by 1,4-diaminobutane or spermidine[12, 13]. In animal tissues amines are derived by decarboxylation of ornithine, rather than by decarboxylation of arginine[14, 15]. Purification of ornithine decarboxylase has been hindered by the very low enzyme activity normally present. However, as discussed in the section on the role of polyamines in growth, ornithine decarboxylase levels increase dramatically after a variety of stimuli, and the enzyme has been purified to apparent homogeneity from regenerating liver[17] and from the livers of rats treated with thioacetamide[18]. Pyridoxal phosphate appears to be a required cofactor[16, 19]. |
|
Physiological function |
Putrescine fulfills important physiological functions in a wide variety of living cells. This BA shows many physiological functions and it is a precursor in the synthesis of other polyamines (spermine and spermidine). Putrescine is classified as a physiologic amine. Physiological functions of putrescine and other polyamines are related to their polycationic nature, which determines interactions with negatively charged molecules such as DNA, RNA, proteins, phospholipids[20]. Newer studies show that putrescine, along with other polyamines and phosphate ions, forms nuclear aggregates of polyamines in the cell nuclei, which are responsible for the abovementioned interactions and affect the 3-dimensional structure of DNA[21]. These interactions are related to the regulation of the structure of nucleic acids and protein synthesis[22, 23]. Putrescine, along with other polyamines, binds to membrane structures such as phospholipids, mainly in erythrocytes. This polyamine may lead to a decrease in membrane fluidity but also to increased resistance to fragmentation due to stabilization of the membrane skeleton[24, 25]. It has also been found that apart from the membrane stabilization and the effect on the synthesis of nucleic acids and proteins, polyamines are involved in the removal of free radicals[26]. In many mammals, they play an important role as luminal growth factors for intestinal maturation and growth[27, 28] and can play a significant role in the prevention of food allergies[29]. In mammals, polyamines have direct effects on several ion channels and receptors, resulting in the regulation of Ca2+, Na+, and K+ homeostasis[30, 31]. |
|
Toxicity |
With respect to important physiological functions, it is clear that disruption of the normal balance due to increased intake of putrescine from food can have serious toxicological consequences. Although the toxic effects of putrescine are significantly lower than that of histamine or tyramine, there are many serious secondary effects. Diamines such as putrescine have a very important role in alimentary poisoning as they can enhance and potentiate the toxic effect of histamine, tyramine, and phenylethylamine by interacting with enzymes that metabolize these BAs[33]. For example, experiments on guinea pigs and rats revealed that putrescine potentiates histamine toxicity up to 10 times[34, 35]. Putrescine enhances histamine toxicity by inhibiting enzymes oxidizing histamine diaminooxidase(DAO; EC 1.4.3.6)?and histamine N-methyltransferase (NMT; EC 2.1.1.8)[36-38]. Moreover, from a toxicological point of view, a serious aspect of putrescine occurrence in foodstuffs is the possibility of forming carcinogenic nitrosamines. Putrescine can form carcinogenic nitrosamines by the reaction with nitrites[39, 40]. |
|
The effect of putrescine on food quality |
In addition to the toxic effects, the occurrence of putrescine in foodstuffs leads to undesirable organoleptic properties and adversely affects the taste and aroma of food[41], for example, in shrimps, it is perceptible at concentrations of 3 mg/kg[42]. Increased occurrence of putrescine indicates food spoilage caused by microbial activity and it is also the main BA that indicates spoiled meat. The amount of putrescine, histamine, and cadaverine shows the freshness of meat and is defined as biogenic amines index (BAI)[43]. |
|
Reference |
Smith TA. 1981. Amines in food. Food Chem 6[3]:169–200. Bard′ocz S, Duguid TJ, Brown DS, Grant G, Pusztai A, White A, Ralph A. 1995. The importance of dietary polyamines in cell regeneration and growth. Br J Nutr 73[6]:819–28. Guggenheim, M. 1 9 5 1 . Die biogenell Amine. Switzerland: Karger. 4th ed. Tabor, H., Tabor, C. W. 1964. Pharmacal. Rev. 1 6:245-300 Tabor, H., Tabor, C. W. 1972. Adv. Enzymol. 36:203-68 Morris, D. R., Fillingame, R. H. 1974. Ann. Rev. Biochem. 43:303-25 Morris, D. R., Pardee, A. B. 1966. J. Bio. Chem. 241:3 129-35 Morris, D. R., Koffron, K. L. 1969. J. Bio. Chem. 244:6094-99 Applebaum, D. 1972. Purification and characterization of induced and biosynthetic ornithine decarboxylases of Escherichia coli. PhD thesis. Univ. Washington. Seattle. 157 pp. Univ. Microfilms #73-1 3789 Wu, W. H., Morris, D. R. 1973. J. Bio. Chem. 248: 1687-95, 1696-99 Holtta, E., Janne, J., Pispa, J. 1972. Biochem. Biophys. Res. Commun. 47: 1165-71 Tabor, H., Tabor. C. W. 1969. J. Bio. Chem. 244:2286-92 Morris, D. R., Wu, W. H., Applebaum, D., Kofi'ron, K. L. 1970. See Ref. 8, pp. 968-76 Russell, D. H., cd. 1973. Polyamines in Normal and Neoplastic Growth, New York: Raven. 429 pp. Herbst, E. J., Bachrach, U.,cds. 1970. Ann. NY Acad. Sci. 171:693-1009 Pegg. A. E . . Williams-Ashman. H. G. 1968. Biochem. J 108: 533-39 Friedman, S. J., Halpern, K. Y., Canellakis, E. S. 1972. Biochim. Biophys. Acta 261:181-87 Ono, M., Inoue, H., Suzuki, F., Takeda, Y. 1972. Biochim. Biophys. Acta 284:285-97 Raina, A., Jiinne, J. 1968. Acta Chem. Scand. 22 :2375-77 Igarashi K, Kashiwagi K. 2010. Modulation of cellular function by polyamines. Intl J Biochem Cell Biol 42[1]:39–51. Di Luccia A, Picariello G, Iacomino G, Formisano A, Paduano L, D’Agostino L. 2009. The in vitro nuclear aggregates of polyamines. FEBS J 276[8]:2324–35. Silla Santos MH. 1996. Biogenic amines: their importance in foods. Intl J Food Microbiol 29[2–3]:213–31. Hou MH, Lin SB, Yuann JM, Lin WC, Wang AH, Kan Ls L. 2001. Effects of polyamines on the thermal stability and formation kinetics of DNA duplexes with abnormal structure. Nucleic Acids Res 29[24]:5121–8. Til HP, Falke HE, Prinsen MK, Willems MI. 1997. Acute and subacute toxicity of tyramine, spermidine, spermine, putrescine and cadaverine in rats. Food Chem Toxicol 35[3–4]:337–48. Largue E, Sabater-Molina M, Zamora S. 2007. Biological significance of dietary polyamines. Nutrition 23[1]:87–95. Kaur-Sawhney R, Tiburcio AF, Altabella T, Galston AW. 2003. Polyamines in plants: an overview. J Cell Mol Biol 2:1–12. Dufour C, Dandrifosse G, Forget P, Vermesse F, Romain N, Lepoint P. 1988. Spermine and spermidine induce intestinal maturation in the rat. Gastroenterology 95[1]:112–6. L¨oser C. 2000. Polyamines in human and animal milk. Br J Nutr 84[Suppl1]:S55–8 Dandrifosse G, Peulen O, El Khefif N, Deloyer P, Dandrifosse AC, Grandfils C. 2000. Are milk polyamines preventive agents against food allergy? Proc Nutr Soc 59[1]:81–6. Dandrifosse G, Peulen O, El Khefif N, Deloyer P, Dandrifosse AC, Grandfils C. 2000. Are milk polyamines preventive agents against food allergy? Proc Nutr Soc 59[1]:81–6. Johnson TD. 1996. Modulation of channel function by polyamines. Trends Pharmacol Sci 17[1]:22–7. Li J, Doyle KM, Tatlisumak T. 2007. Polyamines in the brain: distribution, biological interactions, and their potential therapeutic role in brain ischemia. Curr Med Chem 14[17]:1807– https://www.alfa.com/zh-cn/catalog/B21316/ Taylor SL. 1985a. Histamine food poisoning: toxicology and clinical aspects. Crit Rev Toxicol 17[2]:91–128. Parrot J, Nicot G. 1966. Pharmacology of histamine. In: Eichler O, Farah S, editors. Handbook of experimental pharmacology. Heidelberg: Springer-Verlag. p 148–61. Lehane L, Olley J. 2000. Histamine fish poisoning revisited. Intl J Food Microbiol 58[1–2]:1–37. Stratton JE, Hutkins RW, Taylor SL. 1991. Biogenic amines in cheese and other fermented foods: a review. J Food Prot 54[6]:460–70. Hern′andez-Jover T, Izquierdo-Pulido M, Veciana-Nogues MT, Marine-Font A, Vidal-Carou MC. 1997. Biogenic amine and polyamine contents in meat and meat products. J Agric Food Chem 45[6]:2098–102. Emborg J, Dalgaard P. 2006. Formation of histamine and biogenic amines in cold-smoked tuna: an investigation of psychrotolerant bacteria from samples implicated in cases of histamine fish poisoning. J Food Prot 69[4]:897–906. Shalaby AR. 1996. Significance of biogenic amines to food safety and human health. Food Res Intl 29[7]:675–90. Bover-Cid S, Holzapfel WH. 1999. Improved screening procedure for biogenic amine production by lactic acid bacteria. Intl J Food Microbiol 53[1]:33–41. Lehane L, Olley J. 2000. Histamine fish poisoning revisited. Intl J Food Microbiol 58[1–2]:1–37. Benner Jr RA, Staruszkiewicz WF, Rogers PL, Otwelle WS. 2003. Evaluation of putrescine, cadaverine, and indole as chemical indicators of decomposition in penaeid shrimp. J Food Sci 68[7]:2178–85. Karmas E. 1981. Biogenic amines as indicators of sea food freshness. Lebensmitt Wissensch Technol Food Sci Technol 14[5]:273–5. |
|
Safety Profile |
Poison by subcutaneous, intravenous, and rectal routes. Moderately toxic by ingestion. An experimental teratogen. Human mutation data reported. When heated to decomposition it emits toxic fumes of NOx. See also 1,3-BUTANEDIAMINE and AMINES. |
|
Overview |
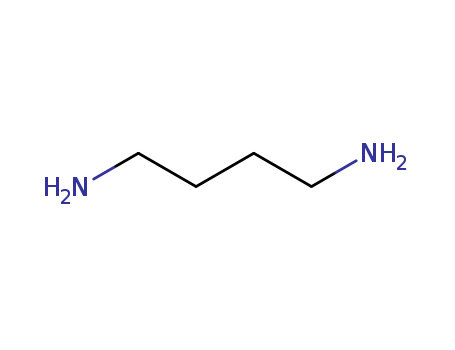
Putrescine is a low-molecular-weight nitrogenous base with the systematic name 1,4-diaminobutane. It is an aliphatic diamine belonging to the group of biogenic amines (Bas). Two basic amino groups are present, which at the physiological pH of 7.4 carry a positive charge that makes them suitable for a wide range of functions in different cell types. According to some authors, putrescine also belongs, together with cadaverine, spermine, and spermidine, to polyamines (molecules containing two?or more amino groups in the molecule)[1,2]. Polyamines are found in all cell types and their presence in various kinds of foodstuffs is partly due to their endogenous origin. In humans, there are 3 common sources of putrescine: the first one is endogenous biosynthesis within their own cells, the second one includes foodstuffs (alimentary intake), and the last one is the production of putrescine by bacteria of the intestinal microflora. The largest amount of putrescine in humans is taken from food[2]. If none of the 3 sources of putrescine becomes excessive, putrescine is used for its physiological functions and the excess is excreted by normal metabolism. However, increased intake of putrescine in food can lead to serious toxicological consequences. Figure 1 The chemical structure of putrescine |
|
Applications |
Putrescine is used as a precursor in many biological systems and synthon for amido-ureas. It is involved in the synthesis of nylon 46 by reacting with adipic acid[32]. |
|
Definition |
ChEBI: A four-carbon alkane-alpha,omega-diamine. It is obtained by the breakdown of amino acids and is responsible for the foul odour of putrefying flesh. |
InChI:InChI=1/C4H12N2/c5-3-1-2-4-6/h1-6H2
Selenomonas ruminantium synthesizes cada...
Lysine decarboxylase (LDC; EC 4.1.1.18) ...
A putrescine bisamide with a unique cycl...
Diaminopimelate decarboxylase (DAPDC) an...
Several NiO-MgO systems were synthesized...
Nickel(II) complexes of 1,2-butanediamin...
Putrescine, spermidine and spermine of h...
Agmatine is a precursor for polyamine bi...
-
Acrolein, a toxic unsaturated aldehyde g...
-
A rhodium-catalyzed method for the hydro...
Sophisticated chemical processes widely ...
The first total synthesis of kukoamine B...
The invention relates to a method for pr...
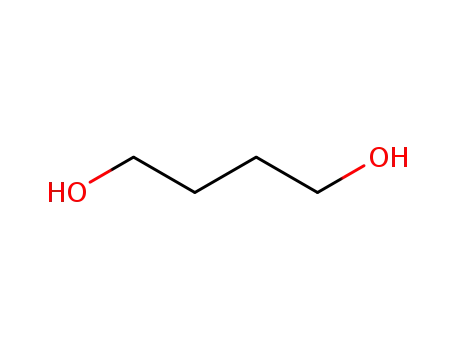
Butane-1,4-diol

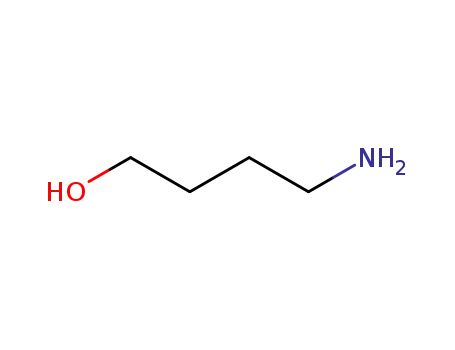
4-Aminobutanol

1,4-diaminobutane
| Conditions | Yield |
|---|---|
|
With
ammonia;
chlorocarbonylhydrido[4,5-bis(dicyclohexylphosphinomethyl)acridine]ruthenium(II);
In
toluene;
at 155 ℃;
for 12h;
under 30753.1 Torr;
Product distribution / selectivity;
Autoclave;
|
|
|
With
ammonia; hydrogen;
chlorocarbonylhydrido[4,5-bis(dicyclohexylphosphinomethyl)acridine]ruthenium(II);
In
toluene;
at 180 ℃;
for 12h;
under 52505.3 Torr;
Product distribution / selectivity;
Cooling;
Autoclave;
|
|
|
With
ammonia;
chlorocarbonylhydrido[4,5-bis(dicyclohexylphosphinomethyl)acridine]ruthenium(II);
In
toluene;
at 155 ℃;
for 12h;
under 30753.1 Torr;
Product distribution / selectivity;
Autoclave;
|
Butane-1,4-diol
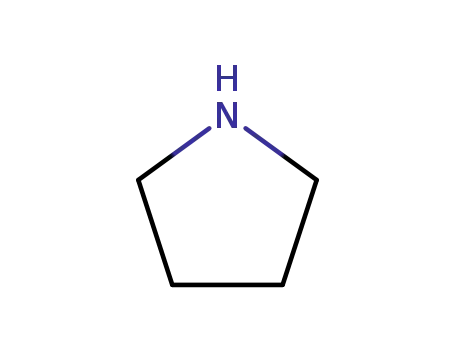
pyrrolidine
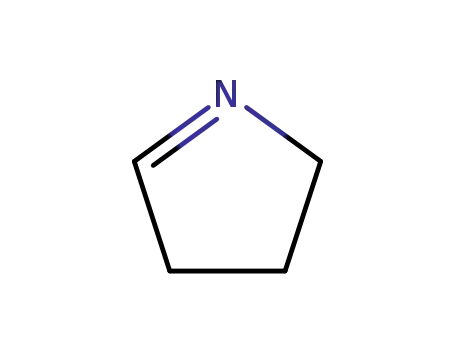
1-pyrroline
4-Aminobutanol
1,4-diaminobutane
| Conditions | Yield |
|---|---|
|
With
ammonia;
chlorocarbonylhydrido[4,5-bis(dicyclohexylphosphinomethyl)acridine]ruthenium(II);
In
para-xylene;
at 180 ℃;
for 12h;
under 38253.8 Torr;
Product distribution / selectivity;
Autoclave;
Inert atmosphere;
|
|
|
With
ammonia;
chlorocarbonylhydrido[4,5-bis(dicyclohexylphosphinomethyl)acridine]ruthenium(II);
In
toluene;
at 155 ℃;
for 24h;
under 38253.8 Torr;
Product distribution / selectivity;
Autoclave;
Inert atmosphere;
|
|
|
With
chlorocarbonylhydrido[4,5-bis(dicyclohexylphosphinomethyl)acridine]ruthenium(II); ammonia;
at 180 ℃;
for 12h;
Temperature;
Inert atmosphere;
Autoclave;
|
|
|
With
chlorocarbonylhydrido[4,5-bis(dicyclohexylphosphinomethyl)acridine]ruthenium(II); ammonia;
In
toluene;
at 155 ℃;
for 24h;
under 30153 Torr;
Pressure;
Reagent/catalyst;
Temperature;
Catalytic behavior;
Autoclave;
Inert atmosphere;
|
|
|
With
chlorocarbonylhydrido[4,5-bis(dicyclohexylphosphinomethyl)acridine]ruthenium(II); ammonia;
In
toluene;
at 155 ℃;
for 12h;
under 31503.2 Torr;
Inert atmosphere;
Autoclave;
|
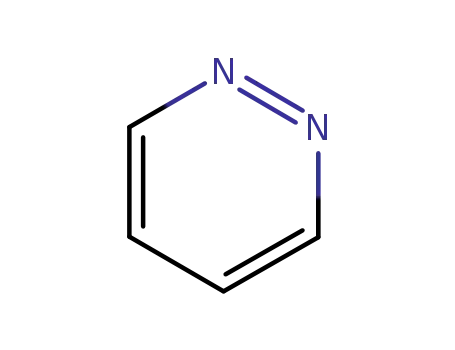
1,2-diazine
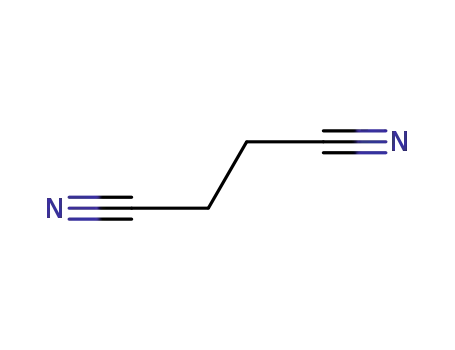
butanedinitrile
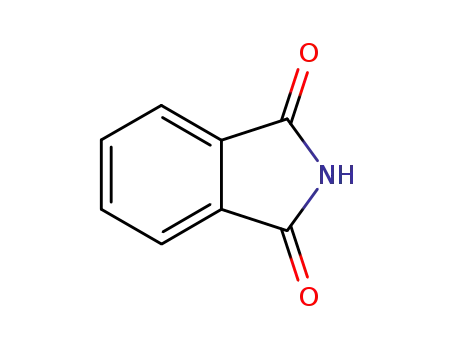
phthalimide
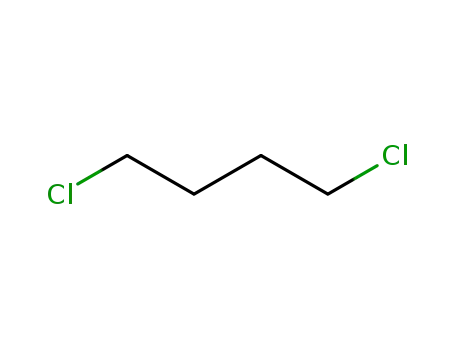
1,4-dichlorobutane
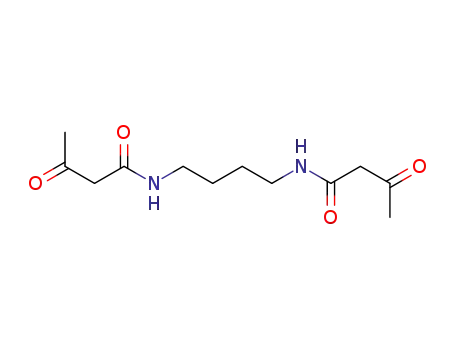
N,N'-tetramethylenebis(3-oxobutanamide)
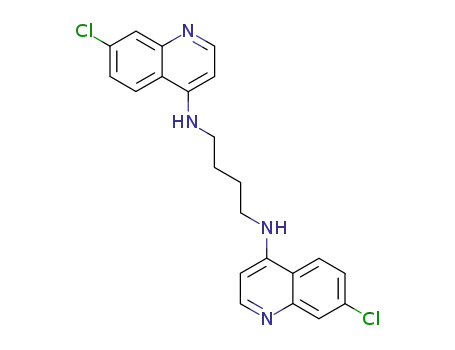
N1,N2-bis(7-chloroquinolin-4-yl)butane-1,4-diamine
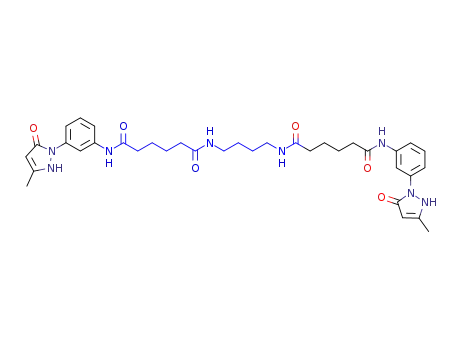
1,4-bis-{5-[3-(3-methyl-5-oxo-2,5-dihydro-pyrazol-1-yl)-phenylcarbamoyl]-valerylamino}-butane
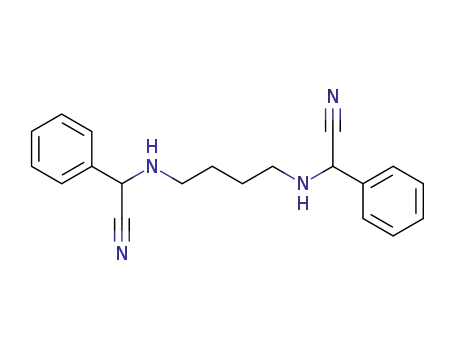
2,2'-diphenyl-2,2'-butanediyldiamino-di-acetonitrile